Our latest work on engineering extrachromosomal circular DNAs (ecDNAs) is now available in bioRxiv. In this preprint we describe a novel strategy to engineer the formation of large (>1 Mbp), multigenic, amplifications mediated by ecDNAs in cells and mice.
ecDNAs are large, circular DNA fragments that mediate focal amplifications in some of the most aggressive human cancers. ecDNA were described more than 60 years ago and they are known as “double minutes” for their appearance in metaphase spreads (see image). ecDNAs replicate once per cell cycle, exactly like normal chromosomes, but because they lack centromeres, they segregate randomly at mitosis. This in turn allows for their rapid accumulation (or loss), depending on whether extra copies of the gene(s) they contain can increase cellular fitness.
The basic idea at the core of our work is straightforward and takes advantage of the ability of the Cre recombinase to induce not only excision, but also circularization of any genomic regions flanked by 2 loxP sites having the same orientation.
Using CRISPR-Cas9, we insert two ‘circularization cassettes’ so that they flank the region we want to amplify. The cassettes contain one loxP site each and are designed so that Cre-mediated recombination generates an ecDNA consisting of the region of interest plus reconstituted GFP and HygromicinR transgenes. In addition, a second fluorescent reporter, mScarlet, is also activated upon recombination, but remains linked to the chromosome.
This design allows us to track cells that have undergone Cre-mediated circularization and monitor ecDNAs abundance over time and in response to different selective pressures.
To showcase this strategy, we engineered the formation of ecDNAs containing the MDM2 oncogene in HCT116 cells, which are otherwise diploid and chromosomally stable.
In this first set of experiments we show that these cells readily form ecDNAs upon Cre-recombination and that we can monitor the number of ecDNAs and their change over time using flow cytometry or microscopy. We can even force accumulation of ecDNAS by taking advantage of the presence of the hygromycing resistance gene in the ecDNAs.
In a second set of experiments, we used this basic strategy to generate two novel genetically engineered mice harboring Cre-inducible ecDNAs containing the Myc or (1.7Mbp) or the Mdm2 oncogenes (1.3 Mbp), respectively.
We show that the engineered Myc-containing ecDNAs provide a strong fitness advantage to primary neuronal stem cells rapidly accumulate to dozens of copies per cell, with some cells having more than 150 copies of the ecDNAs.
The Mdm2-containing ecDNAs also rapidly accumulate in MEFs, where they suppress the p53 pathway, overcome senescence, and promote cancer formation in cooperation with Ras, generating sarcomas that are reminiscent of dedifferentiate liposarcomas (a tumor type that is characterized by the presence of MDM2 amplification in humans).
This is a novel direction for our lab, the result of more than 3 years of work from a group of fantastic students, postdocs, and research technicians. A shout out goes to @prade90, @minsizhang, @ruirui_gao, and
@melissyao, but it was truly a team effort with all members of our lab giving important contributions. Finally, a big thank you to Jens Luebeck and Vineet Bafna, from UCSD, who used their AmpliconArchitect suite to analyze whole-genome sequencing data and reconstruct the ecDNAs in silico.
There are a ton of questions that the tools we have generated will help address. Just a few: can ecDNAs initiate tumor formation in vivo? How do they contribute to tumor progression and metastasis? What are the key requirements for their replication and propagation in primary cells and in cancers? Does their presence create unique therapeutically actionable vulnerabilities? Can they transfer from cell to cell?
Postdoctoral positions to tackle these and other exciting questions are available in our lab.
Congratulations to our own Rui Gao, who graduated last week brilliantly defending her thesis on engineering ecDNAs in cells and in animals!
Rui has been an outstanding member of our lab and we will all miss her very much. She should be proud for having launched several new lines of research that will keep us busy for years to come. Rumors are that she is moving to Washington DC to start a new and exciting professional career. We look forward to learn about her many successes!!!
We are thrilled to share a new paper describing a novel approach to interfere with global microRNA activity in cells and in living animals without affecting miRNA biogenesis or miRNA abundance. You can read the paper on eLife or on bioRxiv, where it was initially posted as a preprint.
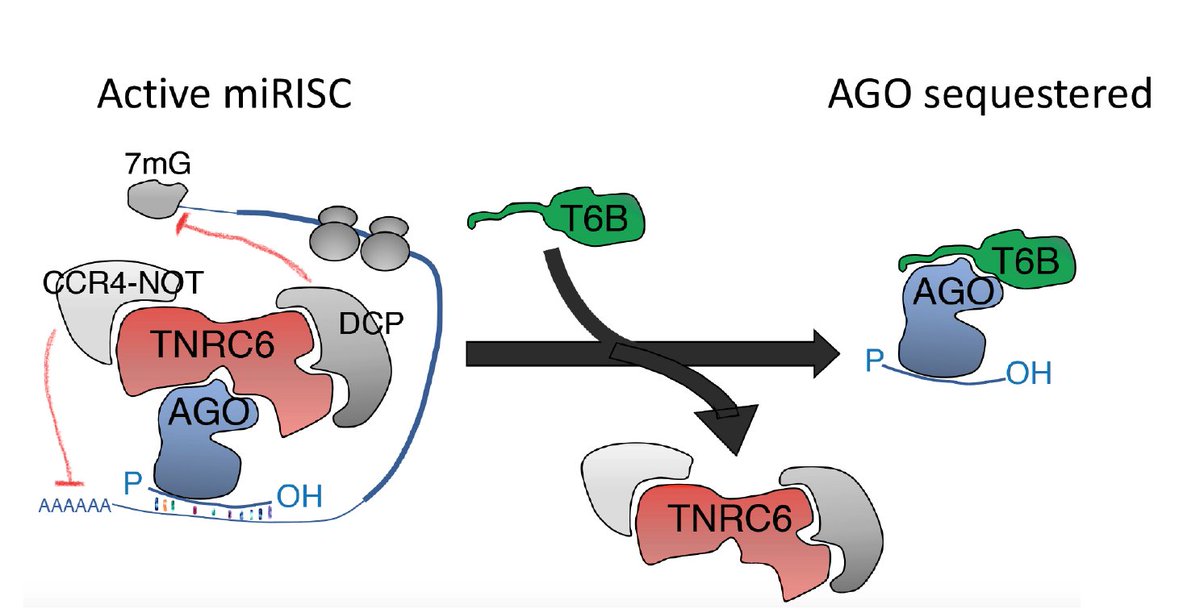
At the core of this work is a peptide (T6B) previously generated by Gunter Meister and Tom Tuschl that binds to Argonaute proteins and prevents the assembly of a functional miRISC complex, thus effectively blocking miRNA-mediated gene repression without impairing miRNA biogenesis.
Schematics of how T6B works
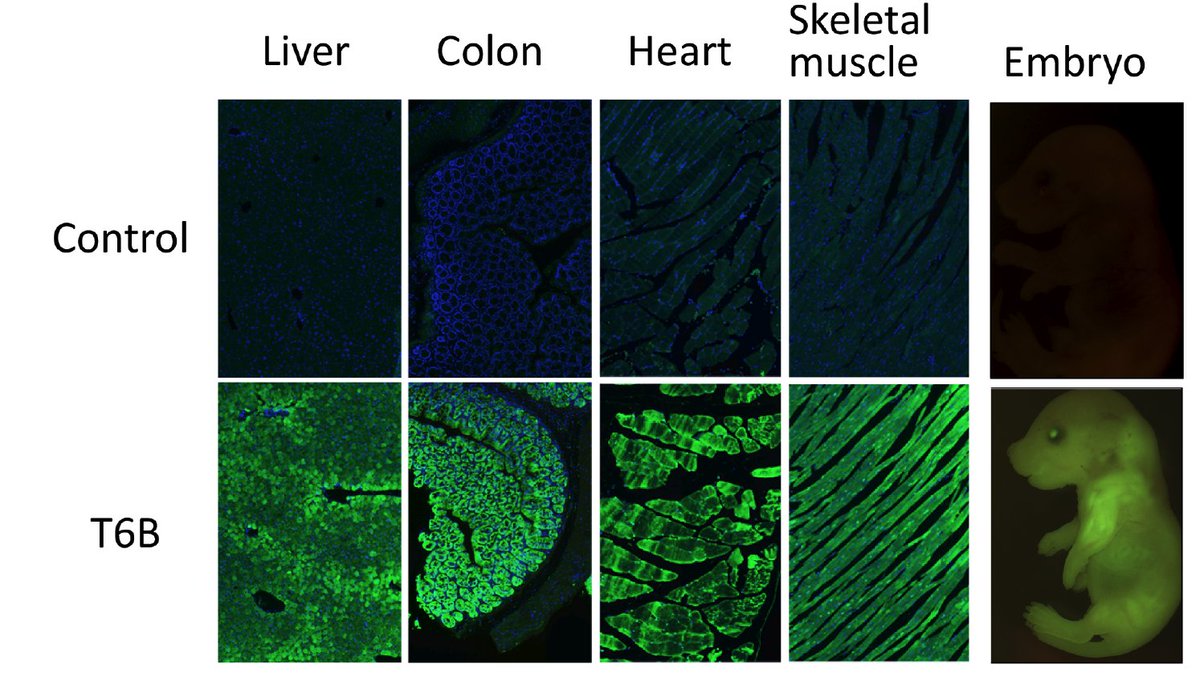
To explore the consequences of miRNA-inhibition in vivo, we have generated a novel genetically engineered mouse strain that expresses the T6B peptide fused to YFP in a doxycycline-inducible manner. By crossing it with an appropriate rtTA expressing strain it is possible to block miRNA function in a tissue-restricted and temporally controlled manner.
T6B-YFP expression in a panel of tissues and in embryos
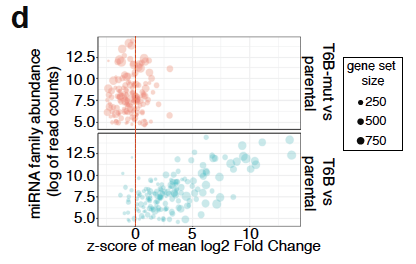
The effect is fully reversible, and miRNA biogenesis (or endogenous siRNA function) are not affected. The entity of de-repression of predicted miRNA targets is roughly proportional to the corresponding miRNA family abundance.
Bubble plot showing global inhibition of miRNA function in MEFs
Each “bubble” in the plot above is a set of predicted targets of a miRNA family. On the x axis we plotted the average log2 fold change compared to the background(reported as z-score) and on the y-axis is the miRNA family abundance. Using this novel mouse strain we show that although miRNA-mediated repression is required during embryonic development, in many adult tissues it seems largely dispensable under homeostatic conditions, and becomes essential only in after acute damage and during regeneration.
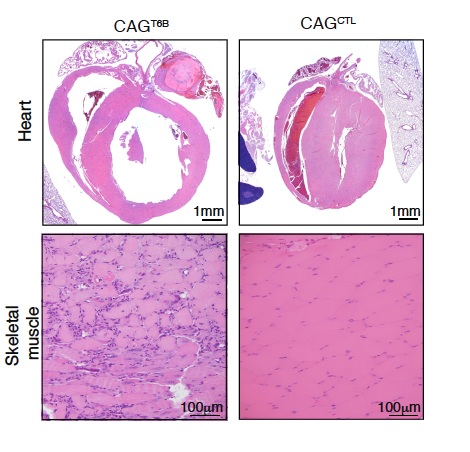
A notable exception is represented by the skeletal muscle and the heart, where even under physiologic conditions T6B induction results in tissue degeneration and death.
Effect of T6B expression on muscle and heart
We have a ton of follow-up experiments planned and we hope our colleagues interested in the biology of miRNAs will find it useful. As always, the mouse strain will be freely available through @jacksonlab as soon as we have completed the donation process.
This work was the result of a collaborative effort between our lab, the Thompson lab, the @KevinHaigisLab lab, the Tuschl lab, the @dbetel lab, the @VidigalJoana lab, and the @MeisterLab.
Dr. Gaspare La Rocca deserves a special mention as he was the main driver of this project. He came up with the idea, performed—together with Bryan King—most of the key experiments, and persisted through many ups and downs during the past few years.
A warm welcome to Davide Pradella, our new postdoc!
Davide is from Gravedona, a small village on the north shores of Lake Como, in Italy. He obtained his Ph.D. in Genetics, Molecular, and Cellular Biology from the University of Pavia, working on RNA metabolism at the Institute of Molecular Genetics (IGM).
Davide research interests will focus on ecDNAs, cancer, and gene fusions. Stay tuned!
A big congratulations to our own Xiaoyi Li who today brilliantly earned her PhD with a wonderful presentation to cap five years of hard work and a remarkable first author paper in Molecular Cell. Xiaoyi will soon move to the west coast to start her postdoctoral training.
Running a lab can be frustrating at times, but the privilege of witnessing a young student like Xiaoyi flourish and blossom into an accomplished scientist is, at the end of the day, what makes our job the best in the world.
Ad majora Xiaoyi, the future is yours! And thanks again for choosing us five years ago…
We have an NIH-funded postdoctoral position available immediately to study BRAF-driven pediatric brain cancers. You will use ex vivo and in vivo somatic chromosomal engineering to model and characterize pediatric brain tumors driven by the BRAF oncogene, with a particular focus on tumors driven by BRAF gene fusions.
The goals include defining the molecular mechanisms driving these tumors and responsible for their unique biological and clinical features, and developing and testing novel therapeutic strategies.
Contact us for additional details and check out our work on CRISPR-based chromosomal engineering.
Our HEAP paper is finally out in Molecular Cell! The result of a close collaboration with Christina Leslie and her group, this work was led by Xiaoyi Li, Yuri Pritykin, and Carla Concepcion. We invite you to read the full paper for all the details, but if you are interested just in the punch line, here is a broad overview.
Xiaoyi, Yuri, and Carla: the brains and the hands behind this work.
MicroRNAs (miRNAs) are evolutionarily conserved small RNAs that mediate post-transcriptional gene repression. They exert the repressive functions through binding to targets, which, in most cases, are the 3’ untranslated regions (3’UTRs) of mRNAs. This interaction is mediated by the miRNA-induced silencing complex (miRISC). The core component of miRISC is the Argonaute (Ago) family protein, which consists of four protein members in mammalian genomes (Ago1-4). Ago2 is the most important member among all Ago proteins (Figure 1). Our group, along with many others, has shown the important roles that miRNAs play under both physiological and pathological conditions 1-3.
Figure 1. The miRISC complex
Over the past two decades a major focus in this field has been to assign phenotypes to individual miRNAs and miRNA families and to define their mechanism(s) of action. Mapping the biological targets of miRNAs has become a key aspect in miRNA research, and a wide range of computational and experimental tools have been developed over the years for miRNA targets determination.
Identification of miRNA targets by crosslinking immunoprecipitation (HITS-CLIP) followed by high-throughput sequencing is the prototype of a class of methods (HITS-CLIP, iCLIP, PAR-CLIP, CLASH, CLEAR-CLIP, eCLIP, etc) 4-9 developed to directly purifying miRNAs and their targets by UV crosslinking and immunoprecipitation of Ago-containing complexes from cells. These methods theoretically provide a transcriptome-wide targeting landscape of all expressed miRNAs in cells.
Although these methods have proven extraordinarily useful in mapping miRNA targets and in learning the rules used by miRNAs to select their targets in various cellular contexts, several intrinsic difficulties have to be overcome before they can be applied to broader cellular contexts, especially, to in vivo contexts. These difficulties include technical complexity and the lack of an in vivo platform which allows flexible targets purification from live tissues/organs.
The motivation of this study was to design a novel strategy to address some of the limitations of CLIP-based methods. To this end we took advantage of the HaloTag system developed by Promega, which offers an easy, antibody-free, approach for protein isolation and labeling 10. The HaloTag is a mutated bacterial haloalkane dehalogenase that catalyzes an irreversible covalent bond between itself and its substrate 11,12. By fusing the HaloTag to Ago2 and using synthetic HaloTag ligands conjugated to beads for complex purification, we developed a simplified miRNA target purification pipeline, built on top of the HITS-CLIP protocol, which can free us from the time-consuming procedures seen in conventional CLIP methods (Figure 2). We named this method Halo-enhanced Ago2 pull-down (HEAP). In the paper we show that the enhanced purification stringency and fewer steps result in miRNA target libraries with great depth, resolution and reproducibility.
Figure 2:outline of the HEAP protocol.
To address the need of in vivo miRNA target purification, we generated a novel genetically engineered mouse model harboring a conditional Halo-Ago2 allele. The HaloTag was knocked in front of Ago2 with a “loxP-STOP-IRES-FLAG-loxP” cassette in between. The Halo-Ago2 fusion is therefore expressed in a Cre-regulated manner, adding another layer of flexibility to this system. We demonstrated the usefulness of this system by identifying targets from several different in vivo contexts, including mouse embryos, adult tissues and primary tumors.
Our group has long-lasting interest in studying the biological functions miR-17~92. To determine the direct targets of members in this miRNA cluster, we generated HEAP libraries from E13.5 embryos lacking miR-17~92. By doing that, we identified a large number of binding sites which contained seed matches for miRNAs in miR-17~92 and whose peak signals decreased when ablating miR-17~92 genetically (Figure 3a). Interestingly we found that a small fraction of Ago2 binding sites mapped to long non-coding RNAs (lncRNAs). One interesting example is Cyrano, a lncRNA containing two miR-92 binding sites (Figure 3b). Differential gene expression analysis in miR-92-deficient mice supports the functionality of these miR-92 binding sites.
Figure 3: HEAP libraries from miR-17~92 mutant embryos.
Lastly, we wanted to look into miRNA regulations under pathological conditions. One interesting setting is primary cancers where massive transcriptomic rewiring has occurred during transformation. To compare miRNA regulations in tumors versus normal cells, we induced primary tumors in the conditional Halo-Ago2 mice and expressed Cre to activate Halo-Ago2 expression. We generated HEAP libraries from two tumor types (glioma and non-small cell lung cancer) and from their corresponding tissues of origin. A direct comparison of miRNA binding sites between tumor versus normal tissues illustrated striking differences between the two. For example, in normal cortex, miRNAs such as miR-124 and miR-128 had large number of targets involved in normal brain physiology, while in tumors, miR-219 became highly “active”, showing the largest number of targets (Figure 4a). We also showed that miRNA abundance was the most likely determinant of miRNA targets seen in each context (Figure 4b). Cross-context comparisons also highlighted the enrichment of miR-17~92 binding sites in both glioma and lung cancers, pointing to a potential general requirement of miR-17~92 for oncogenesis.
Figure 4. HEAP libraries from gliomas and normal cortices.
One aspect we think is important to emphasize is that the Halo-Ago2 mouse strain can be easily employed in conjunction with any of the many already available CLIP variants (PAR-CLIP, eCLIP, iCLIP, CLASH, etc.) to obtain even more granular information on miRNA targets. Finally, the HaloTag provides also an excellent opportunity to identify novel Ago2 interactors and post-translational modifications in vivo, and to image the dynamic of miRISC in living cells.
Video 1, Halo-Ago2 localization in living cells (in collaboration with Ryan Schreiner).
1 Ventura, A. et al. Targeted deletion reveals essential and overlapping functions of the miR-17 through 92 family of miRNA clusters. Cell132, 875-886, doi:10.1016/j.cell.2008.02.019 (2008).
2 Han, Y. C. et al. An allelic series of miR-17 approximately 92-mutant mice uncovers functional specialization and cooperation among members of a microRNA polycistron. Nat Genet47, 766-775, doi:10.1038/ng.3321 (2015).
3 Bartel, D. P. Metazoan MicroRNAs. Cell173, 20-51, doi:10.1016/j.cell.2018.03.006 (2018).
4 Chi, S. W., Zang, J. B., Mele, A. & Darnell, R. B. Argonaute HITS-CLIP decodes microRNA-mRNA interaction maps. Nature460, 479-486, doi:10.1038/nature08170 (2009).
5 Hafner, M. et al. Transcriptome-wide identification of RNA-binding protein and microRNA target sites by PAR-CLIP. Cell141, 129-141, doi:10.1016/j.cell.2010.03.009 (2010).
6 Helwak, A., Kudla, G., Dudnakova, T. & Tollervey, D. Mapping the human miRNA interactome by CLASH reveals frequent noncanonical binding. Cell153, 654-665, doi:10.1016/j.cell.2013.03.043 (2013).
7 Konig, J. et al. iCLIP reveals the function of hnRNP particles in splicing at individual nucleotide resolution. Nat Struct Mol Biol17, 909-915, doi:10.1038/nsmb.1838 (2010).
8 Moore, M. J. et al. miRNA-target chimeras reveal miRNA 3′-end pairing as a major determinant of Argonaute target specificity. Nat Commun6, 8864, doi:10.1038/ncomms9864 (2015).
9 Van Nostrand, E. L. et al. Robust transcriptome-wide discovery of RNA-binding protein binding sites with enhanced CLIP (eCLIP). Nat Methods13, 508-514, doi:10.1038/nmeth.3810 (2016).
10 Gu, J. et al. GoldCLIP: Gel-omitted Ligation-dependent CLIP. Genomics, Proteomics & Bioinformatics16, 136-143, doi:https://doi.org/10.1016/j.gpb.2018.04.003 (2018).
11 Encell, L. P. et al. Development of a dehalogenase-based protein fusion tag capable of rapid, selective and covalent attachment to customizable ligands. Curr Chem Genomics6, 55-71, doi:10.2174/1875397301206010055 (2012).
12 Los, G. V. et al. HaloTag: a novel protein labeling technology for cell imaging and protein analysis. ACS Chem Biol3, 373-382, doi:10.1021/cb800025k (2008).
This year we headed upstate for a two days of sport, science, brainstorming, and good food.
Gaspare entertained everybody with his unsuspected skills as piano player and singer.
The brainstorming session produced many interesting new ideas, with Nathan, Ram, and Jinny securing the top three places (and an Amazon gift card each!).
This slideshow requires JavaScript.
It was also an opportunity to celebrate the first 10 years of our lab, hoping that the next 10 years will be as fun and productive!
We are happy to announce that our former postdoc Joana Vidigal is starting her own lab at NCI. Given her successes as a grad student and as a postdoc it is easy to predict she will be an outstanding group leader. If you are interested in RNA biology and genome editing, her lab is the place to be. For more information, visit Joana’s website.
Ping Mu, our very first graduate student, after completing a very successful postdoc with Charles Sawyers, has now started his own lab at UTSW where he will continue his ground breaking work on prostate cancer. If you are looking for a postdoctoral position I cannot think of a better environment and a better lab! To learn more visit his lab website or contact Ping via email